


Hermaphroditism and Sex Change in Caridean Shrimps
|
|
|
| Rhynchocinetes uritai male phase, example of a purely protandric species (from Bauer and Thiel, 2011; photo by Martin Thiel) | A female-phase protandric individual from a species (Thor manningi) with a mix of protandric and primary male individuals (Bauer, 1986) | Two female-phase simultaneous hermaphrodites (far left, far right) with a male-phase individual in between; this species, Lysmata wurdemanni, is one of several protandric simultaneous hermaphroditic species (Bauer and Holt; 1998; Bauer, 2000) |
Most shrimps have separate sexes (gonochorism), male and female. However, a number of shrimp species are hermaphroditic, i.e. function as male and female sometime during their life cycle. Hermaphroditism may be sequential, with the individual developing sexually first as one sex and then changing to the other. Female to male sex change is known as protogyny, found in many marine fish species. All hermaphroditic shrimps are protandric, first functioning as male and then changing their external and internal sexual characters to female. In recent years, a number of these species have been shown to be protandric simultaneous hermaphrodites(PSH) (Bauer, 2000), first developing as male but when older and larger, developing female characters and reproductive ability while retaining male function as well.
In shrimps, the direction of hermaphroditism can be explained by the energetic difference between eggs and sperm. Egg cells must contain much more energy and mass in order to support embryonic development, while the same energy content can produce many more sperm cells in the males. In many, perhaps the majority of shrimp species, males are much or somewhat smaller than females, which must grow larger in order to produce the much less abundant yolk-filled egg cells. Thus, protandric species usually evolve in shrimp taxa with small males and larger females, a situation common in caridean shrimps. Indeed, hermaphroditic species have only been found found in the Caridea. Paradoxically, penaeoid species are usually have smaller males than females, yet no hermaphroditic species have been found in that group.
Of the 3800 species of carideans, only 90 protandric species have been reported (Bauer, 2023 book). However, the number of shrimp species whose sexual systems have been well studied numbers is in the hundreds, not thousands. But this is changing: in a 2000 review paper, I reported just over 30 protandric species. That number has increased to 90 and there are certainly many more waiting to be discovered.
The first well studied examples of protandric shrimps came from the north Pacific and Atlantic pandalid shrimps important in commercial fisheries. My studies on hermaphroditic shrimps began with a tropical seagrass caridean, Thor manningi (Bauer, 1986). I found that smaller individuals were mainly two types of males, one (primary males) with exaggerated sexual appendices (appendices masculinae = AM) on the pleopods (abdominal swimmerets) and a pair of prehensile legs, while the other male morphotype had much smaller AM. With increasing size, these latter individuals lost male characteristics with a change of testes to ovaries; at a certain size, they lost all maleness and were typical females that incubated embryos. In other words, these "males" are protandric hermaphrodites. In another Thor species, T. amboinensis, all individuals are strictly hermaphroditic (Baeza. J.A. and Piantoni C. 2010. Sexual system, sex ratio, and group living in the shrimp Thor amboinensis (De Man): relevance to resource-monopolization and sex-allocation theories. Biol. Bull. 219: 151165. ).
In 1998, two publications confirmed a new type of hermaphroditic system in shrimps in the genus Lysmata, protandric hermaphroditic hermaphroditism (PSH) (Fiedler, 1998: L. amboinensis; Bauer and Holt, 1998: L. wurdemanni). Since that time, nearly 90 PSH species have been confirmed, mainly Lysmata spp., many due to the efforts of J. Antonio Baeza, a former student in my lab who is now a professor in Biological Sciences at Clemson University. A similar number of protandric species have been identified in the Pandalidae. Protandric species have also been found in the Alpheidae, Atyidae, Campylonotidae, Crangonidae, Hippolytidae, Merguiidae, and Rhynchocinetidae.
A paradox presented by Lysmata species is that all species of the genus studied are protandric simultaneous hermaphrodites. Lysmata species generally fall into two very different groups in their ecological and social characteristics. Many species are primarily nocturnal species with somewhat inconspicuous coloration of reddish stripes or blotches, such as L. wurdemanni and L. californiensis (photo below; Bauer and Newman, 2004). They occur in small to large aggregations and do engage in fish cleaning behavior (feeding on mucus, parasites, dead skin of fishes or other invertebrates). However, cleaning behavior is only occasional and just one mode of feeding. They do not maintain cleaning stations or solicit fishes for cleaning opportunities. Other species, such as L. debelius (photo, below), L. amboinensis, and L. grabhami live in hermaphrodite pairs, mate with each other both as male and female, are active during the day, and maintain cleaning "stations". They are vividly colored, with conspicuous bright white antennae and legs. They attract fishes to their station on a coral head or other reef structure by lashing their conspicuous antennae and swaying back and forth.The fishes approach submissively and allow the shrimps to clean their bodies of parasites, dead skin, and excess mucus. In doing so, the shrimps crawl over the fish's body, among the gills and even within the gaping mouth of larger fishes. As with other cleaner shrimps, even predatory fishes do not eat or harass the shrimp during cleaning behavior. One might assume that related species would show similar ecology and behavior, but this is not the case with these two socioecological groups of Lysmata. A good explanation for this phenonmenon is still lacking (see Bauer, 2007 and Baeza J.A.2013.Multi-locus molecular phylogeny of broken-back shrimps (genus Lysmata and allies): a test of the Tomlinson-Ghiselin' hypothesis explaining the evolution of simultaneous hermaphroditism. Mol Evol Phylogen 69:4662).
 |
 |
| Lysmata californiensis, a nocturnal aggregating PSH species and facultative (occasional) fish cleaner | Lysmata debelius, a diurnal pair-living PSH species that is a dedicated fish cleaner which displays to fish clients, which approach to be cleaned by the shrimps. |
When study the mating system of Rhynchocinetes uritai at the Shimoda Marine Lab in Japan, Martin Thiel and I found that it is a protandric species (Bauer and Thiel, 2011). A then graduate student, now Dr. Yumiko Osawa, did her master's thesis on the life history of this species. Based on monthly sampling, Dr. Osawa found that most individuals were recruited into the shallow subtidal during the late summer and fall, matured the following spring and summer as males, overwintered and then the next spring changed sex to female for their second year of life (Osawa et al., 2015; see below ).
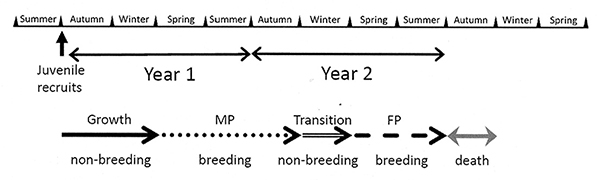
In the genus Rhynchocinetes and the related Cinetorhychus, there are some species with small males and larger females and other with dominant males much larger than females (see Mating Systems and Behavior). We have examined only one small male species (R. uritai) and one large male species (R. typus); the former is hermaphroditic and the latter is not, as expected from the size frequency distributions of the two species. But do the other small male and large male species of these genera follow this pattern? This question, as in other questions about why hermaphroditism evolves in some shrimps and not in others, can only be answered by studies on more species.
Back to Home Page.