Overview: This chapter is arranged in four major sections. The first presents the background to Classical Conditioning, covering Pavlov's work, what he found, what he thought was going on, and what some of the standard paradigms for Classical Conditioning are. The second presents many of the basic principles that determine when conditioning will occur, and whether it will be excitatory or inhibitory. The third discusses important exceptions to these principles, and examines the complex interactions that can arise from the presence of multiple stimuli during various phases of acquisition. Finally, the fourth section briefly examines several alternative accounts of the type of an association that can form, and introduces Rescorla's definition of Classical Conditioning.
Please note the significance of the information in that last paragraph: Many philosophers and scientists had been arguing over a mechanism by which learning could occur, and indeed, over what complex mechanisms would need to be present in order to allow learning. With his interpretation of what he had seen, Pavlov provided an extraordinarily simple and powerful answer to the question of what learning required, and when it would occur. Essentially, all that was needed (well, not really all: We'll get to that in a moment!) was an organism born with certain innate reflexes (see Chapter 1), and the ability to form associations. Stimuli that are present when a reflex goes off will become associated with that reflex. Thus, novel stimuli can come to control reflexes that were previously triggered by a small class of unlearned, innately determined triggers. Or in other words, the ability to associate coupled with the presence of reflexes means that learning will occur. In this fashion, we solve an important mystery of how an organism born with just reflexes and instincts can come to have learned responses.
Pavlov made a number of early discoveries, but the two biggest findings had to do with what he called experimental excitation and experimental extinction. Excitation refers to the ability of a novel stimulus to trigger a response. In a by-now-legendary experiment, Pavlov paired a sound (the ticking of a metronome; the ringing of a bell: a neutral stimulus that by itself does not cause salivation) with the placement of food in a dog's mouth. After several such pairings, the sound acquired the ability to excite drooling. But Pavlov also discovered that when he stopped pairing the sound with the food, the response to the sound eventually went away. He termed this second finding extinction.
We can present models for excitation and extinction. These models will require introducing four technical terms. The terms are unconditioned stimulus (UCS or US), unconditioned response (UCR or UR), conditioned stimulus (CS; but this is a mistranslation from the original Russian that appears to have resisted attempts at correction: the original phrase in Russian is more like conditional stimulus), and conditioned response (CR). The word unconditioned here simply means unlearned. Thus, the unconditioned stimulus and the unconditioned response are the two components of an unlearned, inborn reflex: the trigger, and its result. We can diagram the reflex in this fashion (the arrow here indicates the direction of causality: The stimulus triggers the response):
UCS -----------> UCR
But what about the conditioned stimulus? The conditioned stimulus is a stimulus that is neutral with respect to the unconditioned response. That is, on its own, it doesn't cause that particular response to go off. At the same time, a conditioned stimulus isn't really a neutral stimulus; it often causes an orienting response on the animal's part, because it is a novel stimulus, and organisms, as you know, tend to pay attention to new things. Presumably, for classical conditioning to occur, we need to pair the neutral stimulus with the UCS, like so:
UCS -----------> UCR
+
CS -----------> ???
I won't always use the plus sign in the future (most people don't); I will assume that you understand that the CS and the UCS occur at about the same time when I list them underneath one another in these diagrams. In any case, whatever response the CS triggers at this early stage in the game, it has nothing to do with the UCR; it doesn't resemble it in any way. But, with repeated pairings, the CS comes to take on a response that is normally similar to the UCR. And because of that similarity, we can now redraw the diagram in the following fashion to indicate that learning has been successful:
UCS -----------> UCR
CS -----------> CR
And that is the procedure by which Pavlov obtained excitation. As for extinction, if, after you have already verified that you have a CR, you present the CS by itself a number of times (that is, without the presence of the UCS), then you will find that the CR tends to get weaker until, finally, it is no longer there. So, ringing a bell repeatedly (for our dog who has learned to drool to the bell) will result in cessation of drooling, if the bell and the food are no longer paired.
In the diagram above, we have gone from an unlearned reflex (the UCS triggering the UCR) to a learned reflex (the CS triggering the CR). And unlike most unlearned reflexes (though see the discussion of habituation and sensitization later in this chapter), learned reflexes can apparently go away
As we have seen in the first chapter, Watson was strongly influenced by Pavlov. However, it is worth pointing out that Pavlov was technically not a behaviorist in Watson's mold. One of the major assumptions of Watsonian behaviorism was peripheralism. This was an assumption Pavlov never bought in to. On the contrary, he was a centralist who assumed that much of learning took place in the brain (in the cortical regions). In his account, the stimulus centers in the brain that became active at about the same time would eventually connect together. Or in more modern parlance, borrowing Donald Hebb's famous phrase, neurons that fire together wire together (but don't confuse Pavlov's stimulus centers with Hebb's neurons; I'm providing this phrase as an easy way for you to remember what Pavlov's theory claimed). So, for Pavlov, conditioning occurred because the CS and UCS got connected. His basic idea was the activation or excitation of the CS center in the brain would result in the activation spreading to the UCS center, which would then activate the response.
If you think about Pavlov's theory a bit, you will realize that there are a number of predictions that may be made from it. One important one is that the CR and the UCR should strongly resemble one another; they ought to be the same type of response. The reason is that the CS is really setting off the UCR by means of activating its trigger. Another is that the CR should normally be weaker than the UCR. This prediction arises from the claim that activation or excitation weakens over distance. Thus, the activation that reaches the UCS from the CS is already weaker, resulting in a weaker trigger. (Activation, by the way, is a modern term that resembles Pavlov's term excitation; I'm introducing it here, because it will prove important in later chapters on attention and memory.) A third prediction that may be generated from these is that the strength of the response will depend on the strength of the stimulus (and here we include both the CS and the UCS). The reason is that a stronger stimulus should result in more activation reaching the response center, which, in turn, should trigger a stronger response. A fourth prediction we may wish to add to this cluster is a prediction of directionality. Perhaps the links that are formed in the brain between stimulus centers are directional. On this account, if the CS is presented before a UCS, then the CS center will create a pathway to the UCS center, and activation will flow from the CS to the UCS (I will adopt informal terminology here to simplify the writing: activation doesn't flow between stimuli, but between their centers). Alternatively, if the UCS is presented before the CS, then activation should flow from the UCS to the CS. (This should remind you of Hume's principle of causality discussed in the previous chapter!) Our prediction then becomes that a CR should occur only if the CS is presented before the UCS during training. Finally, there is a prediction of temporal contiguity. The longer the delay between when the CS and the UCS are activated, the less likely they are to wire together. Thus, learning should require relatively short intervals between the CS and the UCS.
How accurate are these predictions?
Pavlov's research did indeed generally support them: CRs resemble UCRs,
but are weaker; more intense CSs and UCSs seem to result in stronger CRs;
presenting the CS first results in an excitatory conditioned response,
but presenting the UCS first generally does not; and long intervals between
the CS and the UCS (by long intervals, we mean measured in seconds!) do
not seem to be effective. Thus, a very simple idea is capable of generating
a number of apparently correct predictions. (Don't take Pavlov's physiology
too seriously, however!)
In Pavlov's view, generalization
was due to a spill-over of excitation or activation, from nearby stimulus
centers in the brain to the CS center (thus triggering it to some extent,
thus causing a flow of activation to the UCS center, thus triggering the
UCR to some extent). He thought that the brain was organized on a principle
of similarity such that similar stimuli would have centers near one another.
(Note how this can account for Aristotle's and Hume's Law of Similarity).
So, fire-engine red should partially activate cherry red,
reddish orange, reddish purple, orange, and purple,
but not blue or butterfly or loud. Moreover, because
reddish orange is closer to fire-engine red than orange
is, presenting reddish orange results in greater activation
of fire-engine red than presenting orange does, thus yielding
a larger CR.
But if you want your animal
to drool to a red stimulus but not to an orange one, how do you turn off
the generalization to orange? Pavlov's procedure here was quite simple:
During training, present both the red and the orange stimuli, but always
pair the red stimulus with the UCS and never pair the orange stimulus with
it. This is the procedure called discrimination; its purpose is
to teach animals or humans to give different responses to two things they
would normally give similar responses to (i.e., to discriminate
between two similar stimuli, or to treat two things differently). The CS
that is supposed to excite the CR (the red light, in this example) is termed
the CS+; and the CS that is supposed to result in no or a very weakened
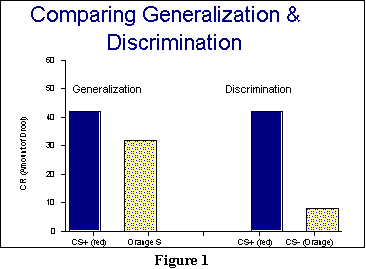
CR (the orange light, in this example) is termed the CS-. Figure
1 presents an idea of what discrimination should accomplish. The two dark
bars (blue, if you have a color monitor) in this figure represent the amount
of drooling we get after training an animal with red as the CS+. The light
(yellow) speckled bars, in contrast, represent the orange stimulus. On
the left, in the Generalization panel, you can see that there is a strong
response to the orange stimulus due to generalization, even though
the animal has received no previous experience with it. But on the
right, where we assume Discrimination Training (which is why the stimulus
is here labeled as a CS-), there ought to be a significantly reduced response.
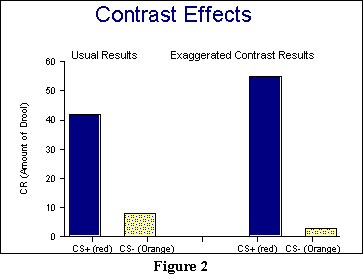
One of the findings Pavlov obtained with the discrimination procedure involves what are called contrasts. What happens in a contrast, in general, is that being exposed to several different things results in exaggerating their differences, at least so far as the responses are concerned. After running through a discrimination procedure like that above, we can obtain a fairly reliable estimate of the CRs to both the CS+ and the CS- by measuring amount of saliva. (If discrimination training has been successful, of course, we should see a much smaller CR for the CS- then in a second group of animals that had only excitation conditioning with the CS+.) Pavlov discovered an additional result, though: If you present a series of trials (after discrimination training) with just one of these CSs, then when you present the other, you will get an exaggerated response to it. So, presenting the CS- over and over again results in an exaggeratedly large amount of drooling when the CS+ is finally presented, and presenting the CS+ over and over again results in an exaggeratedly small amount of drooling when the CS- is finally presented. To give you a handle on what a contrast is like, consider the following example, which is somewhat similar to what we've discussed: The longer you are in a hot room, the colder a cold room will feel when you move into it; similarly, the longer you are in a cold room, the hotter a hot room will feel. Using our example from Figure 1 above, Figure 2 shows you what a contrast might look like. On the left are the normal results to CS+ and CS-, and on the right are the exaggerated results.
Is there evidence in favor of a learning-account of extinction rather than a forgetting account? The answer appears to be yes. There are a number of findings that fit the learning account view. Three that may be mentioned here involve spontaneous recovery, disinhibition, and relearning. Spontaneous recovery refers to an interesting finding regarding extinction: If you wait a bit after extinction training has been completed (so that the animal is no longer exhibiting the response), presenting the CS will once again yield the CR (though only for a few trials). Within reason, the longer the delay after extinction training, the greater the spontaneous recovery. Thus, experimenters who wish to permanently extinguish a response use a procedure of repeated sessions of extinction. The spontaneous recovery after each additional session decreases, until there is finally no spontaneous recovery. With respect to the issue at hand, however, spontaneous recovery should be impossible if the connection or link between the CS and the UCS has dissolved or otherwise truly been forgotten.
Disinhibition also refers to a post-extinction finding. Specifically, immediately following extinction training when the CR no longer occurs, the presentation of a fairly intense novel stimulus seems to temporarily get rid of extinction. One possible explanation of this result (not the only one, of course) is that the new stimulus momentarily increases the animal's attention to its surroundings (to check for danger), including once again attending to the CS. An alternative, nearly opposite theory is that if suppressing the response takes attention, then the momentary disruption of attention ought to temporarily get rid of the suppression: The new situation confuses the animal enough that it 'forgets' to do what it is supposed to do.
Finally, long after extinction, at a time when no spontaneous recovery is present and there appears to be no evidence that the animal remembers the response, we can try to retrain it. The idea behind relearning is that if anything of the original learning has been left over in memory, then retraining should be faster. This finding normally occurs. In fact, we measure the amount of memory involved through what is termed a savings score: the proportion of time saved in learning the second time around. The savings score is:
100 x [ (First Learning - Second Learning) / First Learning ]
As an example, if it took you four years to achieve a certain proficiency in French, and 30 years later, after not having spoken any French, you take courses again but only need two years to acquire the same proficiency, then your savings would be:
100 x [ (4 years - 2 years) / 4 years ] = 100 x (2 / 4) = 50
This is treated as a percentage: Your savings is 50%. And since you learned French in half the time the second time around, you can see that this is what the savings score is signaling. A savings score of 0%, of course, would indicate no advantage of having previously learning the materials: First and second learning would be identical in this case (100 times 0/4 is 0). Thus, the underlying assumption is that any memory left over should give you a non-zero savings.
Savings scores do provide suggestive evidence of memory after extinction, but we need to add several caveats or cautions here. First, looking at relearning immediately after extinction is not appropriate. If the animal is actively inhibiting the CR immediately after extinction, then relearning should take longer (as you will see later, there is some evidence that this indeed happens), resulting in a negative savings score. Second, non-zero savings can also occur because humans and animals are simply getting better at the general ability to learn (what Harlow terms learning-to-learn: see the later chapter on attention). For example, you as a college student should be a better, more sophisticated learner than you were in seventh grade. Comparing your learning now with what you did in seventh grade might thus not be valid.
I've spent a fair amount of time on the notion of inhibition not only because it is important in its own stead, but also because it plays a role in the previous findings on contrasts, and in the next set of findings I want to discuss. Thus, in contrasts, the exaggeratedly positive response to the CS+ (termed positive induction) is presumably due to greater than normal excitation having been triggered; the exaggeratedly negative response to the CS- (termed negative induction) is presumably due to greater than normal inhibition having been triggered (see Figure 2 above). But for Pavlov, inhibition also explained why higher-order conditioning was less successful.
CS Original (CS1) -----------> CR
+
New CS (CS2) ----------->
???
And if second-order conditioning has been successful, then we ought to obtain a conditioned response:
CS Original (CS1) -----------> CR
New CS (CS2) -----------> CR
Moreover, once that link has been established, we can now try for third-order conditioning, by pairing yet another new stimulus with the second learned reflex obtained from second-order conditioning:
2nd-Order CS (CS2) -----------> CR
+
New CS (CS3)
-----------> ???
And so on. What Pavlov discovered in these types of experiment was that each additional order resulted in a weaker CR. He reported that conditioning beyond the 3rd Order was quite difficult to obtain.
Using Pavlov's theory, we may essentially come up with two complementary reasons why higher-order conditioning should be weaker. The first is that we are creating a longer and longer path between the CS we present, and the UCS center. Thus, in 4th Order conditioning, activation has to travel from CS4 to CS3 to CS2 to CS1 to the UCS. We ought not be surprised if relatively little activation reaches the UCS in this case, resulting in only a weak response. The other reason, however, is that in this procedure, we are also presenting a CS without a UCS. That, of course, is the definition of the extinction procedure. Thus, during 2nd Order conditioning, two very different things are going on: One is that we are trying to establish an excitatory link between CS2 and CS1. The other is that the link between CS1 and the UCS is becoming inhibitory. So, a weaker response may be due to actually training some aspects of inhibition. (We will return to this claim later, to see how accurate it is).
That gives a brief introduction
to Pavlov's theory, and some of the major findings and principles he reported.
Since Pavlov's time, of course, the field has exploded with findings, methods,
and theories. Before going on to the next section, we will close out this
section by briefly introducing some of the major classical conditioning
paradigms. These include excitatory-appetitive conditioning; excitatory-aversive
conditioning, inhibitory conditioning, compound conditioning,
and learned taste aversions.
In excitatory-aversive conditioning, on the other hand, the UCS is normally something the animal would seek to escape or avoid; what, in operant conditioning, we will refer to as a punisher. The UCS here might be a shock of some sort. People often think that use of shock ought to result in inhibition rather than excitation, but that is too simple a view. A dog shocked on its paw will flex or move that paw (particularly if the paw has been resting on a metal surface that can be used to transmit a mild shock). If you pair the shock with the ringing of a bell, then the bell comes to excite the paw flex, not inhibit it. But, there is also a component of inhibition in this situation that we can identify. An important point in classical conditioning is that most theorists believe there to be at least two sorts of conditioning going on in any given experiment: the conditioning of a physical response, and the conditioning of an emotional response. Emotionally, a negative UCS should cause negative emotions such as fear or pain or frustration that will interfere with (or inhibit) ongoing activity. So, if the CS becomes an inhibitor in this sense (because it is associated with danger), presentation of this danger signal (to borrow Bolles' terminology) should tend to stop the animal doing some other excitatory response. This involves what is called a suppression ratio: To see how emotionally inhibiting a stimulus is, present it while the animal is engaged in an excitatory response such as pressing a bar for food. The amount of decrease in the excitatory response is your measure of the amount of suppression caused by the inhibitory stimulus. One more point is worth introducing here, although it really comes to us from operant conditioning research: After extinction of an excitatory-appetitive CR, the CS may come to act as an inhibitory stimulus that has a measurable suppression ratio. Again, there are a number of theories that could account for why this happens; for the moment, let us go back to Amsel's Frustration Theory introduced in the previous chapter. A CS signaling something you want is a good thing. But when you get the CS and are expecting food and the food doesn't come, that causes frustration. And that frustration, of course, can become associated with the CS, so that you would prefer to avoid it.
We have already talked to some extent about inhibitory conditioning, especially with respect to extinction. You will see in the next section below several other techniques that can result in inhibition. For now, however, you should know that there are certain phenomena that resemble inhibition, but are not regarded as being true inhibition. Thus, tests have been developed to help identify true inhibition. Most theorists argue that true inhibition requires a certain result on both the summation and the retardation tests. The summation test involves an idea very similar to the suppression ratio mentioned above: If a CS is inhibitory, then it should interfere with excitation, so that another excitatory response weakens in the presence of that CS. If you want, you may think of excitation and inhibition as being generalized positive and negative emotional states that determine how strongly a response is performed. The presence of inhibition takes away from the overall level of excitation, thus resulting in a weaker response. In the retardation test, in contrast, we attempt new excitatory learning with the supposedly inhibitory CS. The notion here is that the learning will first have to overcome the inhibition, so it should take longer. The reason we cannot rely just on the retardation test, for example, is that slower learning may also be due to an animal that is no longer paying attention to the CS, a very different phenomenon from inhibition and its negative connotations (see the next chapter). In one situation, the animal would try to avoid the CS (i.e., when it is inhibitory); in another, the CS would simply be irrelevant to the animal (i.e., when it is no longer attended).
Compound conditioning will prove quite interesting. It is not the case in the real world that only one stimulus is present when a reflex occurs. So, a number of researchers have also done studies that go beyond simple conditioning (where there is one CS paired with the UCS) to situations where multiple CSs may be paired simultaneously with one CS. Schematically, compound conditioning may be represented as follows:
UCS
-----------> UCR
+
CS1 CS2 CS3 ----------->
???
Do all of these stimuli condition equally well with the UCS? Do they all yield conditioned responses? There will turn out to be some surprising answers here.
There will also be
some surprising answers regarding the learned-taste aversions paradigm.
Here, we attempt to classically condition an avoidance to a certain taste.
This paradigm deserves separate discussion because many of the findings
obtained with it will apparently differ from those obtained in other paradigms.
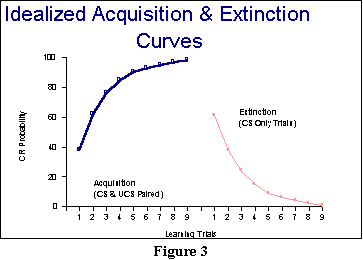
Finally, we should note something
about what learning looks like in classical conditioning. Normally, if
we graph the learning or acquisition curve (in which
we look at how much better learning is becoming with each additional
pairing of the CS and UCS), we find what is called a negatively-accelerated
or diminishing returns curve. Figure 3 (left panel) presents
an idealized example in which we measure learning by how probable
the CR is (though we could have also looked at how strong the CR
was becoming; both are legitimate measures of learning). Negative acceleration
in this curve means that the curve is slowing down; we are getting smaller
and smaller changes as we proceed. And that is also what is meant by diminishing
returns: The biggest changes occur early in learning. Of course, since
extinction is considered to be a type of learning, you would expect an
inverse type of curve, as shown on the right-hand side of Figure 3: With
a few trials of extinction (in which the CS is presented without
the UCS), the curve starts falling off dramatically. With more and more
trials, the changes become smaller in part because there is less change
possible (since you're getting closer to the 0 level). In both learning
and extinction, the portion where the curve seems to be going flat is called
asymptote. In the figure, acquisition and extinction appear to be
symmetric (i.e., the rate of change is approximately the same in each).
In real experiments, they are not. There appear to be several major differences
between excitatory and inhibitory learning, but among the major differences
you should know about is that excitation builds up faster than inhibition.
But how fast does a learning
curve rise? How long does it really take for learning to occur?
The answer to that will depend on a number of things, as you will see.
However, with the right conditions, it is possible for learning
to occur quite rapidly. For example, consider learning in two related excitatory-aversive
paradigms that are often employed: the nictitating membrane response
in rabbits, and the human eyeblink. In the nictitating membrane
response paradigm, a slight electric shock is given to the region surrounding
a rabbit's eye, and this causes a so-called third eyelid (known as the
nictitating membrane) to descend. In the human eyeblink paradigm, a puff
of air is the (unconditioned) stimulus to the eye, which causes a blink
(the unconditioned response). Measuring changes in electrical activity
in the muscles controlling these responses demonstrates that an association
can form in as little as eight or so trials, although more pairings of
the CS and UCS will normally be required to see the full-blown response.
And in cases where a severe aversive UCS is employed, we sometimes
see conditioning in only one or two trials, conditioning so strong
that it resists normal extinction training. In classical conditioning,
the CER paradigm is the name usually given to the method
that employs an aversive stimulus likely to cause a strongly unpleasant
affective state (such as fear), because that method is designed to result
in a conditioned emotional response.
As an alternative, if there is a delay between the CS and the UCS, then you can look for a response that occurs after the CS has been presented, but before the UCS has occurred. In this case, the response is assumed also to be due to the CS, and so, to be a learned response rather than a UCR. If all you look at, however, is whether the response is there or not, then obviously, this procedure will not allow you to claim that a response following the UCS is learned; if the response occurs after the UCS, we have to assume that it is due to the UCS rather than the CS.
Or do we? Another possibility is that we can look at the time line of a response. If you know about how long it takes to get a response from the UCR, then a response that comes after the UCR but appears too quickly to fit within the UCR's normal time line might just be a conditioned response. But this type of procedure, of course, will depend critically on the interval between the CS and UCS: A larger interval gives you more opportunity to see a clean conditioned reponse, but a larger interval also has the disadvantage of resulting in weaker conditioning (see below).
As the discussion demonstrates, there are many issues to be considered in deciding how to measure learning. Among the characteristics of an association that can be used to assess strength of learning, however, four are worth noting. These are a conditioned response's amplitude; its latency; its probability of occurrence; and the effect of a CS's presence in some other task (i.e., presence of interactions).
Amplitude refers to the strength or vigor of a response. Typically, as an association strengthens over repeated pairings, the response should grow stronger. Thus, in the example I gave you above using the nictitating membrane or the human eyeblink paradigm, we can often measure a response early in learning that is so weak that a full eyeblink or a drop of the nictitating membrane has not occurred: We know there is learning because we are picking up changes in signals to the muscles that will be responsible for those movements. And with further pairings, the signals become stronger, to the point that they trigger a response that may now be observed without the need for special instruments. Even so, you can imagine measuring how strong an eyeblink is, and expecting a stronger blink with more pairings of the CS and the UCS. A strong blink should occur in a shorter period of time, for example. And in theory, differences in amplitude of the response might serve to distinguish whether the response is conditioned or unconditioned.
Alternatively, we can measure how long it takes for the response to fire off (not the same as how fast the response takes to execute, once it starts, as in the eyeblink example above). The assumption here is that as learning progresses, resulting in a strengthening association, you should get a CR occurring at smaller delays. Looking at how long it takes before a CR is triggered involves the measurement of the response's latency. Latency, of course, can be used to some extent with both of the general procedures above: Where you intersperse catch trials with training trials, and where you only have training trials (so you look at whether a response occurs before the UCS has been presented).
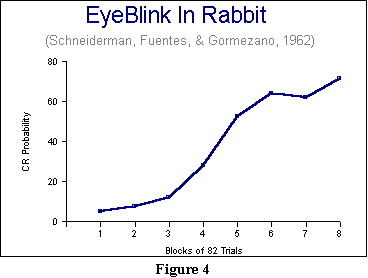 Another measure of associative strength that may also be used with either
the catch-trials or no-catch-trials procedure is to look at probability
of a CR. Here, over a predefined block of trials, we can ask how often
we saw the CR. Figure 4 presents some data by Schneiderman, Fuentes,
and Gormezano, using the eyeblink paradigm with rabbits. They looked
at successive blocks of 82 trials, and simply tracked the proportion of
times a CR occurred between the CS and the UCS. As you can see, even after
656 pairings, the response was occurring less than 75% of the time.
Another measure of associative strength that may also be used with either
the catch-trials or no-catch-trials procedure is to look at probability
of a CR. Here, over a predefined block of trials, we can ask how often
we saw the CR. Figure 4 presents some data by Schneiderman, Fuentes,
and Gormezano, using the eyeblink paradigm with rabbits. They looked
at successive blocks of 82 trials, and simply tracked the proportion of
times a CR occurred between the CS and the UCS. As you can see, even after
656 pairings, the response was occurring less than 75% of the time.
While we have some real data up, for a change, let us also note several features about the learning curve in Figure 4. The first is that it is not quite the same as the idealized curve presented in the previous figure. In fact, it has a sort of S shape in which very little happens at first, and then big changes occur, followed by successively smaller changes. The second feature is that the reason for the discrepancy in real and ideal shapes may have to do with the notion of a criterion. There may be a certain level of excitation or activation of the muscles that is required for a full-blown response to be visible (and thus countable). Until we reach that level, it will look as if very little is occurring. Thus, you should regard the idealized learning curve of Figure 3 as a hypothetical representation of how an association strengthens. Adding in bridging principles to tie association strength to the criterion for responding in a physical system allows us to explain why real curves differ somewhat from the idealized curves. This point will become important in the next chapter, because we will be introduced to a theory that attempts to generate the type of curve found in Figure 3. The third point to notice is there is some suggestion that the curve may be flattening out, or reaching asymptote. In fact, mathematically, we do have enough data here to fit an idealized curve, and see where that curve predicts asymptote to be. I earlier described asymptote as the portion of the curve that goes flat. Technically, asymptote is the final value the curve is approaching. We regard that value as important in classical conditioning, because many theorists believe that value represents the maximum associative strength that can form. More on this in the chapter below.
Since we have talked about response characteristics thus far, let us now briefly turn to stimulus characteristics. It may at first seem strange to look at whether learning has occurred in terms of the CS rather than the CR, but really, we have been doing that all along. Immediately above, we simply asked what kind of a response could be obtained from it, and looked at specifics of that response. Earlier, in speaking about suppression ratios and the summation tests, we were introduced to the idea that a CS can have an influence on a totally different situation. And that becomes relevant here. Even if we do not look for a physical CR, we can check for evidence of excitation or inhibition by seeing whether simple presence of the CS affects an ongoing excitatory or inhibitory response. A CS in excitatory-aversive conditioning should take on some of the characteristics of a danger signal; we would expect it, following the formation of an association, to interfere with an organisms's appetitive behavior. And by the same token (although this situation is a bit more complex, and less to be relied on), a CS that has successfully been conditioned in excitatory-appetitive conditioning (and may thus be regarded as a safety signal of sorts) ought to mitigate or ease an organism's avoidance behavior. In other words, in these two circumstances, the CS should give an animal signals that conflict with the signals of other stimuli in the animal's environment. We ought to see the results of those conflicting signals in terms of moderation of what an animal does. Thus, there should be an interaction of the CS with the other stimuli present.
As this is in part a course in which you ought to gain some appreciation of scientific technique in action rather than in the abstract, let us diverge a moment to discuss methodology. Normally, in an experiment in psychology, the experimenter systematically changes one or more independent variables that are predicted to have an effect on behavior (according to some theory), and measures the result in terms of one or more dependent variables that capture aspects of the supposed effect. As you should recall from other classes, the reason we term possible results dependent is because we are normally testing out an idea that they depend on the presence or level of the independent variable. Thus, in Figure 4 above, the independent variable was the number of blocks of conditioning, and the obvious prediction was that more blocks should result in better learning. The dependent variable that we used to measure amount of learning was the probability of a CR. And what we saw was a systematic correlation between the independent and dependent variables: More blocks resulted in a stronger probability of a CR, as expected. Often, however, our independent variable can involve groups that get different experiences. We will see a number of situations in which there is a control group that receives no special treatment, and an experimental group that gets something special. The question we will subsequently ask is, do we find a difference between the groups (on the dependent variable)? If so, then our working assumption is that the difference is due to the treatment. And if not, then we assume that the treatment is ineffective.
Why bring this up, at this point? The answer is, looking for an interaction can be a risky procedure. A CS can influence on-going appetitive or aversive behavior for reasons that have nothing to do with learning. Whatever an animal is doing, it may be momentarily distracted by the CS, which, of course, will moderate the behavior. So, to do an interaction check properly, we would want to do a follow-up experiment. In our experiment, we will assume that animals have been taught something like bar pressing in the presence of a humming noise. Then experiment will then be something like the following.:
Group Phase 1 Phase 2
1
CS + Aversive UCS
bar pressing with humming noise
2
CS + Aversive UCS
bar pressing with humming noise and CS
3
CS alone
bar pressing with humming noise and CS
Let's go over why each group is here. First, if we compare Group 1 with Group 2, Group 1 acts as the control group to see, in Phase 2, whether presenting the CS causes our animals to change how often they press the bar. So, in Phase 2, the independent variable is presence versus absence of the CS. Presumably, they are pressing the bar for food (excitation). If the CS has become associated with danger, it ought to interfere with bar pressing, slowing it down some. That, so far, is the type of measure of learning I described, several paragraphs above, as looking for an interaction (I am not using this term in its technical statistical sense, for those of you wondering why this wouldn't be a main effect in an analysis of variance: It would!).
But we also need the Group 3 to nail down that interpretation. Thus, Group 3 acts as a control group for Group 2: Groups 2 and 3 are identical in Phase 2, but differ in Phase 1. Specifically, there is no classical conditioning for Group 3 (so the independent variable is presence versus absence of classical conditioning). Thus, if Groups 2 and 3 are identical in Phase 2, then this tells us that the reason Groups 1 and 2 were different had nothing to do with the CS being associated with an aversive UCS! But why on earth would Groups 2 and 3 be identical in Phase 2? Well, several possibilities could account for this. One is that the CS may simply distract the animal, and that is why it bar presses less. Another is that the CS is not neutral, but is itself an aversive stimulus interfering with bar pressing excitation. Running Group 3 lets us know whether we should seriously consider these possibilities. Ideally, we should find that Groups 1 and 3 are identical, so that any difference in Group 2 is specifically due to association of the CS with the UCS.
The moral of the story is
that an effect can sometimes be due to very different causes, including
things we did not think about. That is in part why I stated in Chapter
1 that killer experiments were rare. The usual process involves
a lot of mopping-up experiments designed to prove to others that your interpretation
of why the results occurred is accurate. In order to do that, you need
to consider alternative explanations, and eliminate them. In much of this
work, I will be presenting studies that do have many alternative
explanations. I won't always (or usually) present the mop-up studies, but
you should be aware that they are often there.
In simultaneous conditioning, we directly address the issue of having the CS and the UCS paired at the same time. To keep things simple for the moment, let us assume that each stimulus is presented for exactly the same amount of time. That being the case, the CS and the UCS both come on at the same instant, and some time later, both go off at the same instant. In fact, we might be tempted to make a very strong prediction here. Based on the fire-together, wire-together rule, we might expect this to be the strongest learning of all. Indeed, from everything you've read about so far, this seems a natural prediction; if association is a process of connecting things that have temporal and spatial contiguity, then the greater the contiguity, the better or stronger we would expect the association to be.
So, here is the surprising result: It turns out to be very difficult to get any long-term excitatory learning going on in this situation! Some excitatory responses early in learning are sometimes reported, but they are relatively rare, and they do not last. And that, in turn, may suggest to you that there is perhaps something seriously wrong with a simple temporal contiguity principle. On the other hand, UCSs by definition are significant biological events. They may play such an important role in an animal's life that it has little attention to dispose on other stimuli. Some such explanation could account for the lack of association with simultaneous conditioning.
One more result concerning simultaneous conditioning deserves to be mentioned: There are a sufficient number of studies (not all, however) that report successful emotional conditioning, when the UCS is aversive. That is, presenting a CS with an aversive UCS does seem to imbue the CS with some negative quality, as assessed through suppression ratio or interaction-type experiments. Thus, in part contrary to general process theory, we find some preliminary evidence that associations sometimes form differently for appetitive and aversive UCSs.
Our second paradigm will involve what is variously called forwards or delayed conditioning. In this paradigm (and again, we'll assume equal durations of CS and UCS, to keep the discussion simple), the CS comes before the UCS, but overlaps somewhat with it. At issue is how much earlier the CS is. That, in turn, depends in part on how long the CS and the UCS last, since a duration of 0.2 sec means that the CS has to come less than 0.2 sec before, in order for there to be any physical overlap. (If the durations of both stimuli are 2 sec, in contrast, then the CS can come nearly 2 sec before, and still be on when the UCS starts.) Normally, delayed conditioning yields a fairly strong association, so long as the delay isn't too small (approaching simultaneous conditioning), or too large. Depending on the species and the response, a delay of 0.2 sec to slightly over 0.5 sec serves to give strong conditioning.
We have already briefly mentioned the results found in backwards conditioning when we discussed Pavlov's findings. If forwards conditioning involves the CS coming first, backwards conditioning involves the CS coming second. Generally, backwards conditioning does not result in long-lasting excitation. Under special circumstances, some excitatory CRs may be seen after only a few trials; but training with many trials fails to display a conditioned response. Based on this finding, Pavlov and others have sometimes claimed that there is simply no learning in backwards conditioning. As we will see shortly, this is too strong a statement; there most certainly is a type of learning that occurs with backwards conditioning, if you know what to look for!.
Trace conditioning refers to a procedure that seems to rely more on memory. Trace is the technical term sometimes used for memory. In trace conditioning, we have a situation quite similar to delayed or forwards conditioning with the following difference: The CS ceases before the UCS starts. So, there is no physical overlap between the two. Thus, the UCS is being associated with the memory of the CS. Trace conditioning is certainly successful if the interval is short enough, but most studies that have tried to compare it directly with delayed conditioning claim stronger association when there is a physical overlap. In any case, as an example of trace conditioning, we can consider an experiment by McAllister using the human eyeblink paradigm. McAllister used intervals that ranged from 0.1 sec to 2.5 sec between the CS and the puff of air. There was no CR in the 2.5 sec condition. So, there is some evidence here that some type of nearness in time between the CS and the UCS is important (so long as events are not simultaneous).
In fact, a principle of temporal contiguity does make sense (and we can even come up with a reason why simultaneous conditioning ought to be ineffective: see the discussion of signal value below). Recall that Hume developed a principle of causality. Short intervals make sense for such a principle: Effects tend to immediately follow their causes. So, association may be an adaptive evolutionary mechanism that is sensitive to causality in the real world. On this account, stop and think for a moment about what happens if there is a long delay between a CS and a UCS. Other stimuli and events become increasingly likely to occur in that interval, and thus increasingly likely to capture the animal's attention. A different, irrelevant stimulus may thus become connected with the UCS. Or to put this another way, long intervals increase the number of possible causes, thus decreasing the probability of selecting the right cause. (There is another factor to long delays, but we'll get to that in a moment.)
Trace conditioning isn't the only type of conditioning that relies on memory, however. A final type of conditioning does so as well, and has no physical CS that by itself triggers the CR! This is temporal conditioning. In this paradigm, the UCS is repeated at a certain regular interval. After a while, you will see a CR shortly before the UCS is set to occur. Apparently, the time interval here serves as the CS. That is, the animal knows about when to expect the UCS, and starts reacting accordingly.
Another temporal consideration concerns the interval between trials (that is, the interval that starts when the previous UCS has been presented, and ends when the next CS is presented), This interval is called the inter-trial interval (ITI). As the ITI increases, all other things being equal, the conditioning improves. So, according to this isolation effect (Gibbon, Baldock, Locurto, Gold, & Terrace), isolating the CS and UCS pairing by not having anything else happen for a while makes for better learning. This should make sense to you, since having CS-UCS pairings relatively close together means that a UCS is not only preceded by a CS, but also followed by one (allowing for some confusion in the directionality of the link). This finding may be related to a more general finding in psychology called the spacing effect. The spacing effect refers to what happens when we compare learning or practice under massed practice and spaced practice conditions. In massed practice, the same amount of learning is done in a short time; in spaced practice, there are more and longer breaks. The spacing effect is the finding that spaced practice leads to better learning and performance. This should be of relevance to you, because the spacing effect also applies to how you study for a test: If you devote eight hours to studying, spacing those eight hours out will result in better learning than cramming them all together.
There is more to the isolation effect, however. Interestingly enough, it interacts with the principle of temporal contiguity of CS and UCS. As the ITI increases, there is some evidence to believe that longer and longer CS-UCS intervals may still be effective. So, how long a delay may be tolerated in trace conditioning will depend partly on what the ITI is.
Finally, the relative lengths or durations of the CS and the UCS can also have effects, although the findings here turn out to be quite complicated. But to give you a taste of what these may be like, consider a situation where there is a very long CS followed by a fairly short UCS. In this case, it may be that a CR occurs to the CS going away, rather than to the CS coming on! So, as you will also see in the section below on stimulus intensity, a change of some sort may act as the learned stimulus that now triggers a new response, but that change may involve removal of some physical stimulus. The relevance of this, of course, relates to the implicit Watsonian claims that association occurs because two physical events are present at about the same time.
In fact, there are a number of circumstance besides extinction in which inhibition arises, as verified by the summation and retardation tests. These seem to have in common the opposite of a temporal contiguity principle (though you should be aware that we are not claiming inhibition and excitation are mutually exclusive; in certain situations, aspects of each may be conditioned). Thus, other paradigms in which inhibition arises include trace conditioning with long delays; backwards conditioning; discrimination training; and the explicitly unpaired procedure.
A study by Hinson and Siegel will illustrate inhibition in trace conditioning with long delays. They used a trace conditioning procedure in which the CS and the UCS were separated by 10 sec. The paradigm involved the nictitating membrane response in rabbits. These animals failed to show a CR (the eyeblink) to the CS alone, but in later training and testing, the CS interfered with another excitatory response (the summation test), and exhibited significantly slowed forwards conditioning (the retardation test). Thus, whereas short delays can result in excitatory conditioning, longer delays can result in inhibitory conditioning. The problem is that the CS can come to signal very different things in these two situations. With short delays, the CS may be telling you that the UCS is about to appear. But with long delays, it may be signaling a period of time in which the UCS will be absent. Thus, on this account, learning can involve information about whether or not to prepare for the UCS (a point that will prove important when we discuss the Rescorla-Wagner theory in the next chapter).
Recall that Pavlov and others had claimed that there was no learning in backwards conditioning. The basis of this claim was that no excitatory CR occurred (a claim that isn't always true, as we noted above). However, a number of theorists who have used the summation and retardation tests report that the CS becomes inhibitory here. Following up on the theme we introduced in the previous paragraph, a CS that comes after a UCS may be signaling a period of time in which the UCS is absent. Or to put it slightly differently, there is temporal contiguity between the CS and the absence of the UCS! But note a significant point, here! At any given moment in an animal's life, there are numerous stimuli around that could be CSs, and that occur in the absence of a UCS. Why don't all stimuli become inhibitory unless a UCS is temporally near? What this point illustrates is that inhibition is dependent on a UCS being present or (in the case of extinction) expected. The absence of a UCS has to be noted in order for inhibition to build up.
You will now understand how discrimination training results in inhibition. The CS- is never presented with the UCS, whereas the CS+ is always presented. Since we know from generalization that the CS- would normally receive some unlearned excitation, we have a situation in which inhibition builds to cancel the excitation. This example also illustrates the claim that inhibition is dependent on a UCS being expected.
We spoke of control groups above. In many early experiments on classical conditioning, a control group for learning involved presenting the CS and UCS separately, rather than pairing them together, like so:
Group Phase 1 Phase 2
experimental CS + UCS
test for CR
control
CS, UCS separate
test for CR
The control procedure here
is the explicitly unpaired procedure, and corresponds to what we
do in discrimination training with the UCS and the CS- (though note that
there is no CS+ in the control group). Many researchers thought
it would be a good idea to control for how familiar an animal was with
the CS and the UCS, and that was the purpose of this group. But it turns
out that while there certainly is no excitatory CR in the control group,
we do get inhibition to the CS. Thus, looking for how much learning
occurs when two situations are as nearly identical as possible (in terms
of number of times a CS or a UCS is presented, but differing in whether
CS and UCS are paired) turns out not to be so easy: There is learning in
both situations, although of a different sort!
First, the UCS intensity effect does seem to work as advertised: More intense UCSs do result in stronger CRs. But does a stronger UCS really result in faster learning? The answer to this particular question may be that it seems to, but that it doesn't always. In some instances, if you track the learning curves for several different intensities of the UCS, you will see that the learning curves differ primarily in where the final asymptotes appear to be. Specifically, training with more intense UCSs yields learning curves with higher asymptotes. The higher asymptotes often do translate to more vigorous CRs. But, the learning curves appear to approach their asymptotes at about the same rate. And what this means, in contrast, is that on any trial prior to reaching asymptote, the more intense UCS will appear to have resulted in a stronger response than the less intense UCS. Because the responses differ, we do have the impression of faster learning for the stronger response. But here is the critical point: If we define the learning on any given trial as what proportion of the distance to asymptote has been covered (i.e., what percentage of the total possible learning has occurred), then we will find that the same amount of learning has occurred for the different intensity UCSs. In some sense, people like Rescorla and Wagner claim that UCS intensity determines how strong an association can form (as signaled by where asymptote is).
On the other hand, there have also been studies varying UCS intensity that suggest higher intensities may also influence rate of learning; that is, how fast asymptote is reached. Many of these studies tend to involve excitatory-aversive conditioning. In addition, especially in excitatory-aversive conditioning, the duration of a UCS may have an effect similar to intensity. A less-intense shock UCS that lasts for a relatively long time may be equivalent to a more intense shock UCS that lasts for a relatively short time.
It turns out that somewhat different results are obtained with CS intensity. Changes in CS intensity often do influence the course of learning, helping to determine how rapidly the final asymptote will be reached. And in this respect, it is proper to talk about how the intensity of the CS can alter the speed with which something gets learned (i.e., how quickly the learning curve goes flat). For most practical purposes, different CS intensities will result in different-strength CRs.
We need to add a few more wrinkles here, before finishing our discussion of CS intensity effects. One has to do with contrast effects, and involves a study by Grice and Hunter. Using human subjects, they conditioned the eyeblink response to a soft or a loud sound. The soft sound had an intensity of 50 db, and the loud sound had an intensity of 100 db. They ran three groups of subjects, however. Group 1 received just the 50 db CS, and Group 2 received just the 100 db CS. In this experiment, probability of a CR remained under 100%. If we look at CR probabilities as a measure of strength of the association at the end of the experiment, then we get the results that appear on the left-hand side of Figure 5 (I'm using the dark blue bars for the loud sound, and the yellow speckled bars for the soft sound). So far, this is the CS intensity effect in which the faster learning with a stronger CS gives us a stronger conditioned response.
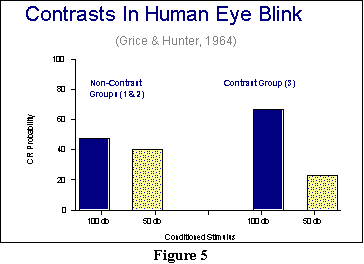 A third group, however, had exposure to both the loud and
soft CSs: One or the other of these was paired with the UCS in random fashion
on any given trial , so that people never knew which CS would appear. The
results for this group are presented on the right-hand side of Figure 5.
And what we see here are the exaggerated results typical of a contrast
effect: positive contrast to the 100 db CS coupled with negative
contrast to the 50 db CS. CS effects thus depend in part on other levels
of the CS to which you have been exposed. You may wish to compare these
results with the positive and negative induction Pavlov
obtained (see Figure 2). Both experiments involved contrast effects, but
with quite different procedures.
A third group, however, had exposure to both the loud and
soft CSs: One or the other of these was paired with the UCS in random fashion
on any given trial , so that people never knew which CS would appear. The
results for this group are presented on the right-hand side of Figure 5.
And what we see here are the exaggerated results typical of a contrast
effect: positive contrast to the 100 db CS coupled with negative
contrast to the 50 db CS. CS effects thus depend in part on other levels
of the CS to which you have been exposed. You may wish to compare these
results with the positive and negative induction Pavlov
obtained (see Figure 2). Both experiments involved contrast effects, but
with quite different procedures.
The other wrinkle involves
a study done by Kamin. Kamin put rats through CER training
using noise as a CS, and a shock as the UCS. (In this experiment, of course,
we use suppression rations to track the success of the conditioning.)
A loud sound was much more effective as a conditioned suppressor
than a soft sound. That fits in with a CS intensity effect. However, the
sounds in this experiment were sudden increases that occurred in a quiet
environment. Kamin was also interested in what might happen if we reverse
the situation: start out with a noisy environment, and use a decrease
in noise level as the CS. So, in a second experiment, the background noise
level of the room was at 80 decibels (db), and the CS was a temporary decrease
(right before the shock) to 70, 60, 50, 45, or 0 db. In this case, the
exact opposite result occurred: The 45 db and 0 db groups displayed
the best conditioning. Since these CSs (particularly in the last
case) involved the smallest physical intensities, a CS intensity
account of the results is no longer possible. Rather, what we appear to
have is a CS salience effect: The more a CS stands out from its
background, the better the conditioning is likely to be. Thus, the important
factor is how noticeable a CS is: Noticeable stimuli are more likely
to be attended or rehearsed, increasing their odds of being connected to
the UCS. And since there was conditioning with a drop to 0 db, we must
again speak of a CS not so much as a physically present stimulus
(see the discussion of temporal factors above), but as some sort of change
that precedes a significant biological event.
As you can see, CS strength
does not seem to involve precisely the same mechanism as accounts for UCS
intensity effects, contrary to Pavlov's view.
Contingency refers to a type of dependency or correlation, whereas contiguity refers to CS and UCS being paired. While the two concepts often make the same predictions, they need not do so. In one famous experiment, Rescorla trained a number of groups of dogs using the CER procedure. When the groups had the same contiguity but differed in contingency (that is, when the CS and UCS were paired the same number of times in each, but additional CSs or UCSs were added in several groups to vary the probability that the CS was a good predictor of the UCS), then the learning changed. This experiment demonstrates that contiguity by itself cannot account for all aspects of learning in classical conditioning.
Rescorla added a further experiment, however, to examine contingency. On his account, learning should only occur when there is some correlation or contingency between the CS and the UCS. Thus, if the CS and UCS are presented at random, even though there will be some accidental pairings of the two (i.e., some contiguity), Rescorla claimed that no learning would occur in this situation. On the other hand, if there is a correlation, then we can ask whether the correlation is positive or negative. In a positive correlation, the CS tends to predict presence of the UCS: When the CS is present, the UCS tends to occur, and when the CS is absent, the UCS tends not to occur.
In a negative correlation, the reverse is true: The CS tends to predict absence of the UCS. Rescorla argued that positive correlations ought to result in excitation; negative correlations ought to result in inhibition; and zero correlations ought to result in no learning.
To study this claim, Rescorla again used the CER paradigm with dogs. He set up an experiment with four groups as follows:
Group CS & UCS Relationship
1
CS + UCS always paired
2
CS, UCS separate (explicitly unpaired procedure)
3
CS, UCS randomly presented (some contiguity)
4
CS, UCS pseudo-randomly presented (some contiguity & contingency)
Groups 1 and 2 are the positive and negative correlation groups in this experiment. Based on what we have already learned, it should not surprise you that we get quite different results out of them. Namely, in Group 1, the CS does indeed act as a conditioned suppressor. However, in Group 2, where the CS predicts absence of the shock, the CS acts as a safety signal that counteracts fear. (This is getting a bit confusing, but recall that an explicitly unpaired procedure should result in inhibition. Since the CER involves excitatory-aversive conditioning, this procedure should result in inhibition of aversion, thus negating a negative emotional condition.)
But what about Groups 3 and 4? They both received random combinations of the CS and the UCS, so that there would be some contiguity that should occur by chance alone (in fact, the same contiguity in each). In Group 4, however, Rescorla added one more clever wrinkle: He got rid of any UCS that came 30 sec or more after a CS. And what that means is that there was also some slight contingency in Group 4. Specifically, 30 seconds is a long enough time that it is a no-CS condition. So here, fewer shocks occurred in the no-CS condition than in the CS-condition, meaning that presence of a CS was a better predictor of upcoming shock. As expected by his contingency account, the CS in Group 4 acted as a conditioned suppressor when he measured its suppression ratio. In contrast, Group 3's CS showed no effect: It was neither a suppressor of excitation, nor a releaser of inhibition. There was no evidence of any conditioning in Group 3. To summarize the results, then:
Group Result & Condition
1
CS ---> conditioned fear (positive contingency)
2
CS ---> conditioned safety (negative contingency)
3
CS remained ineffective (zero contingency; some contiguity)
4
CS ---> conditioned fear (positive contingency; some contiguity)
In the language we have introduced earlier, Group 3 is a control group for Group 4. They have the same contiguity, but differ in whether there is contingency or not (the independent variable). Contingency results in learning, but contiguity by itself does not.
Signal value is inevitably
wrapped up with the problem of selectivity in models like the Rescorla-Wagner
model. That is, speed and success of learning will depend on the animal's
ability to focus in on a good signal. But, a number of factors will influence
the success of that process. Some of these you have already been introduced
to. A noticeable signal, for example, will be checked out sooner than a
less-noticeable signal (the salience effect). Noticeability can
involve strength differences from a background (as discussed above), but
noticeability may involve other factors such as whether a stimulus is presented
by itself, or in the presence of other stimuli. A signal close in time
to the UCS, all other things being equal, will be checked out sooner than
one further removed. A relatively short-lasting signal tends to be better
than a relatively long-lasting one, perhaps because of greater ease in
locating when the UCS should occur following the CS. (Do you expect the
UCS a certain amount of time after onset of the CS? After the CS
stops? After some spot in the middle of the CS? With short
CSs, these issues become less critical.) A signal that has proven successful
in the past may interfere with what actually turns out to be a better signal
in the present (see below). Alternatively, a signal that has been a poor
predictor in the past may no longer be considered in the present, even
though conditions may have changed sufficiently to make it a very good
predictor of the UCS. And all things considered, a signal that has a higher
correlation with the UCS should be better than one that has a lower correlation.
Thus, if one signal successfully predicts the UCS 40% of the time and another
successfully predicts the UCS 67% of the time, we would expect to find
indications in the animal's behavior of the second signal being the more
important. Selectivity implies circumstances in which multiple stimuli
may be present. We consider some of these circumstances and issues in the
next section below.
A study by Obrist, Sutterer, and Howard will serve to illustrate this point. In their study, a CS of a tone was paired with a UCS of shock. One of the reactions to shock (the one we will track as the response) is a heightened heart rate. In fact, in preliminary conditioning, there is evidence that the CR does result in a faster heart rate, perhaps due to the fear that becomes associated with it. However, after a number of pairings, the CS will trigger a response that is a lower than normal heart rate. Thus, the CR appears to become the opposite of the UCR.
This is an interesting finding for a number of reasons. But among them is that this result does a very good job of illustrating the adaptive value of learning (in this case, of classical conditioning). To see why, recall that a CR tends to occur with decreasing latency as learning progresses. Thus, at some point (if your interval between the CS and the UCS allows), you should see the CR after the CS has been presented, but right before the UCS is about to appear. Now add to that the fact that an increase in heart rate is physiologically stressful. In this case, the CS appears to trigger a response that anticipates and attempts to counteract a potentially dangerous increase. An animal receiving shock here will indeed still exhibit an increase in the heart rate as its UCR, but if that animal starts from a lower-than-normal rate, then the increase will bring it more in line with normal levels.
Note several fascinating points here that will be suggestive for some of the theories we look at in the next chapter. If the CR functions to counteract potentially damaging effects of a UCR, then (1) the latency of the CR should depend more on when the UCR would be expected to occur then on strength of the CS-UCS association, and (2) this type of learning only makes sense if a UCR is expected. In other words, the association that is learned is not arbitrary; it presumes that the UCR will be part of the picture. This fits in quite well with theories like the Rescorla-Wagner theory that learning involves animals searching for signals (the CSs) to predict an upcoming UCS (and its UCR), thus allowing them to prepare for that event.
There is some evidence to believe that a similar mechanism accounts for drug tolerance. A drug acts as a UCS that has a certain effect (the UCR). But, the conditions under which the drug is taken (what it looks like; where it was administered; etc.) act as a CS. With antagonistic conditioning, the CS comes to counteract the effects of the drug in advance, so that higher levels are needed to get the same UCR. Thus a vicious cycle is set up requiring greater and greater doses. But the scary part is that if the CS conditions change sufficiently (so that an antagonistic, counteractive CR does not occur), then an overdose becomes possible. People who shoot up in strange or novel environments may not be capable of tolerating the same doses they use in familiar environments (and indeed, there are reports of overdose deaths involving dosage levels that were not unusual for the individual).
Let's take one more example here: a study by Siegel. Siegel used an insulin injection as his UCS. His subjects were rats. The UCR involved a change of blood sugar: a progressive decrease over the course of an hour. But, we can also point to several elements as constituting the CS: the injection; the procedure involved in handling the rats during the injection; perhaps the fact that a blood sample was taken at the same time (blood sugar levels were assessed at various times after the insulin injection to track the reaction). Following training with this procedure, Siegel also injected his animals with a saline solution that would normally have no effect on blood sugar. He obtained an antagonistic response: a higher-than-normal level of blood sugar (which would have helped counteract the low levels caused by the insulin). In this case, the antagonistic conditioning occurred quite rapidly: It was present after only a handful of trials with the insulin injection.
Whatever the exact mechanism
for antagonistic (or as it is sometimes called, compensatory)
conditioning turns out to be, it clearly is inconsistent with a
simple associational model that directly (Hull) or indirectly (Pavlov)
allows a CS to excite the UCR. At the very least, we need to start considering
the possibility that conditioning may be a complex process in which both
excitation and inhibition play a role. Some theories such as opponent
process theory discuss this, and attempt to describe the circumstances
under which one or the other of two opposite processes (excitation; inhibition)
that will both be present will come to dominate the response. In
this theory, consistent with the results obtained by Obrist et al.,
excitation usually builds first, with inhibition kicking in later. Thus,
another suggestion that arises from this work is that excitation and inhibition
are not exactly equal processes; the learning conditions vary from one
to the other.
Group Phase 1 Phase 2
control (Nothing)
CS & UCS paired
1
UCS presented alone CS
& UCS paired
2
CS presented alone
CS & UCS paired
Here, our control group essentially gets to rest and relax during an initial phase of the experiment, while the other two groups are exposed either to various presentations of the UCS or various presentations of the CS. Under these conditions, something quite interesting happens: The learning in Phase 2 for our two experimental groups is impaired. The finding regarding pre-exposing the UCS is called the UCS pre-exposure effect; but the finding regarding the CS is called latent inhibition. Go figure.
Why are these pre-exposure effects problematic for our simple associationistic models (Hull and Pavlov)? The answer is that, in Phase 1, there appears to be no association going on that might interfere with the later attempt to connect the CS and the UCS. Note that we have set up something very similar to the retardation test in Phase 2; we find retardation for both Group 1 and Group 2, compared to our control group. So, does this mean that there is inhibition going on? And if so, how could a CS in Group 1 that never appeared in Phase 1 be inhibited? Why should a CS in Group 2 that had nothing to do with any sort of a UCS be inhibited? As you may perhaps suspect, inhibition will not turn out to be the answer.
In fact, not only will inhibition not turn out to explain these results, but the results themselves will turn out to have different causes. Thus, the UCS pre-exposure effect is believed by many theorists to involve early association-based learning interfering with later learning. That is, despite appearances, many people will claim that the UCS in Group 1's preliminary phase is actually associated with a CS! But of course, this explanation won't fly for Group 2, which is why that group's results are of particular interest.
So what is happening with the UCS pre-exposure effect? Basically, even though there is no explicit CS there, the UCS is normally given in the same context or environment. Elements of that context or environment may then serve as CSs. So, in the absence of a better signal, the environment comes to predict the UCS. And if the animal is using the environment as a predictor, it may subsequently fail to search for another better signal, or it may take longer to realize that there is another better signal. That is why retardation occurs in Phase 2. (You may be wondering why the CS in Phase 2 would be regarded as a better predictor than the environment. The answer is that the CS ought to do a better job of isolating when, in time, the UCS will occur.)
Is there any evidence to support this claim of contextual conditioning? The answer is yes. One piece of evidence that is particularly compelling involves the following type of experimental design:
Group Phase 1 Phase 2
1
UCS alone in Context 1
CS & UCS paired in Context 1
2
UCS alone in Context 2
CS & UCS paired in Context 1
Hinson, among others, has set up this type of experiment (using excitatory-aversive conditioning). The formation of an association (Phase 2) in Group 2 is much stronger. This is exactly what would be predicted from contextual conditioning: For Group 2, the UCS is surprising in Context 1 when they first come across it! They know to expect the UCS in Context 2, but not in Context 1. So, because there is not any sort of a predictor in Context 1, they now look for one. In this case, the CS has good signal value (certainly better than Context 1 cues alone).
One more aspect of the UCS pre-exposure effect is worth mentioning. Particularly with mild shocks in excitatory-aversive conditioning, the UCS pre-exposure effect may in part also be due to habituation. Habituation refers to the finding that animals appear sometimes to 'get used to' a stimulus, in the sense that the response to that stimulus decreases. An orienting reflex to a stimulus tends to habituate on repeated presentations of that stimulus, for example. (Indeed, this is one way in which we study the development of memory in infants: If. For example, we expose very young infants to a face and then later present that face along with a new face for them to look at, memory of that face should result in habituation to it. That is, we should find that they look more at the novel face.) So, if our animal is getting used to mild shocks, then the reason for weak or no responding in Phase 2 (for Group 1) may be that the CS is being associated with a relatively weak UCR. That cannot be the whole story, however, as it would predict a weak response in Group 2, as well. However, if habituation is also dependent on forming an association with the environment, then we may perhaps change our minds about that last prediction. Change the environment, and you also get rid of the cues that tell the animal to habituate.
For many years, habituation was regarded as a non-associative type of learning. Today, some forms of habituation which are relatively long-lasting do appear to be associative. Specifically, there is also evidence that habituation sometimes goes away in different environments. Thus, habituation may be a type of classical conditioning in which the environment acts as a CS. If we add that the CS undergoes some antagonistic or compensatory conditioning (resulting in the opposite of the UCR), then we may perhaps have a mechanism that explains the apparent weakening of a UCR in habituation. (But this is speculation on my part.)
In any case, whether the UCS pre-exposure effect directly involves habituation or not, there is ample evidence for some sort of contextual conditioning going on with pre-exposure.
But what about pre-exposing the CS? There is no UCS present anywhere in this situation, so that it becomes difficult to use a similar account of conditioning. Moreover, it turns out that while the CS in this situation passes the retardation test, it fails to pass the summation test: The apparently "hidden inhibition" (that's what latent inhibition means) that we obtain from pre-exposing the CS is not true inhibition at all!
On the other hand, latent inhibition does share one important feature with the UCS pre-exposure effect, as an experiment by Anderson, O'Farrell, Formica, and Caponegri demonstrates. Anderson et al. used the following design (compare it to the one above!):
Group Phase 1 Phase 2
1
CS alone in Context 1 CS
& UCS paired in Context 1
2
CS alone in Context 1
CS & UCS paired in Context 2
And with this design, they also found that the pre-exposure effect went away with a change in context, arguing for some contextual-based learning. But what is the nature of this learning? In order to account for this type of finding, many theorists now believe that we need to talk about what causes a stimulus to be salient, or distinctive (recall that these are important features of signal value). One of the characteristics of such a stimulus will certainly be that it is associated with relatively unusual events (like a UCS). But, if the stimulus is initially not so associated, then when it is first presented, it will cause an orienting reflex (assuming the stimulus is distinctive enough). That reflex will habituate as the stimulus is repeatedly presented in the initial phase. The stimulus stops being surprising, because it is now part of the background context. As such, the animal no longer pays attention to it. And since it is one of many stimuli in the background context, it is unlikely to be attended to in Phase 2. Or put another way, a new stimulus comes along, you check it out, it doesn't seem to be important, so you stop paying it any heed: it is no longer one of the events you would look to as being important when it comes time to figure out what a good signal for the UCS is.
In this case, the animal
learns about its environment, and what types of things are found in that
environment. That such learning can affect later classical conditioning
is interesting, and suggests that classical conditioning will be influenced
by other types of learning (although learning about what types of stimuli
occur in specific contexts can also be described as S-S or stimulus-stimulus
conditioning, like classical conditioning with its two stimuli).
CS2 -----------> CS1 -----------> UCS -----------> UCR
Here, activation from CS2 flows to the CS1 center, and from there to the UCS center, causing the UCR to go off (which in Pavlov's model was the observed CR). Thus, if we deliberately cause increased inhibition by putting first-order conditioning through a thorough extinction procedure, then we ought to stop that flow, resulting in getting rid of the CR. I'll use diagonal lines to indicate the stopping or inhibiting of that flow. So, what Rizley and Rescorla will attempt to do is see if the following occurs, as predicted by Pavlov:
CS2 -----------> CS1 ----/ /-----> UCS -----------> UCR
If it does, then there is no longer any way for the CS2 to excite the UCR.
They started out by teaching several groups of rats to press a bar for food (this is operant conditioning, of course: see two chapters ahead). Then they set up first-order classical conditioning using a CER paradigm: The UCS was a shock, and the CS was a light. As you know from excitatory-aversive experiments, the light should become a conditioned suppressor under these circumstances. The light, if turned on while the rats are bar pressing, should slow them down! And since this is an experiment on higher-order conditioning, they followed this up by pairing the light with a second CS. Next, they extinguished first-order conditioning in one group of rats by presenting the light by itself ( a second group didn't have the extinction procedure). So, the question is whether CS2 is an equally good conditioned suppressor for both groups. Schematically, the design is this:
Group Phase 1 Phase 2
1 extinction of 1st
Order check for CS2
suppression of bar pressing
2 (Nothing)
check for CS2 suppression of bar pressing
Based on Pavlov's claim, Group 2 should display some suppression for the second CS due to higher-order conditioning. But Group 1 should not: Since the negative qualities that were associated with the first CS were extinguished in Group 1, CS2 should no longer be effective.
So what did they find? I have left out one more wrinkle here that has to do with the fact that there were really two Group 1s! They differed in what CS2 was. For Group 1a, a tone was used (so second-order conditioning involved pairing the light with a tone). For Group 1b, however, another light was used: second-order conditioning of two lights. And what they found was that whether second-order conditioning extinguished along with first-order conditioning differed in these two groups. In the group with the two lights, higher-order conditioning was affected by first-order extinction, as predicted by Pavlov's model. But this did not happen for the group with the tone. Thus, there was some strong evidence against Pavlov: When second-order conditioning involves a change in sensory modality, it apparently can become separated from first-order conditioning.
Other studies have obtained compatible results. Sometimes extinction of first-order conditioning affects second-order conditioning (especially when the modalities of the two conditioned stimuli are the same), and at other times it doesn't. The first result is consistent with Pavlov, but the second isn't. Moreover, it's also the case that the strength of the CR in higher-order conditioning co-varies with the strength or salience of the first-order CS, and with the strength of the UCS. These results are also consistent with Pavlov. If you form an association between CS1 and CS2, then a stronger CS1 should result in better second-order conditioning, just as a stronger UCS results in better first-order conditioning. But the fact that there is also a result of UCS strength suggests an alternative possibility: Maybe CS2 associates directly with the UCS. In order for it to do that, the presence of CS1 would have to remind the animal of the UCS, so that it forms an association between CS2 and the mental representation of the UCS.
In short, some evidence is
consistent with Pavlov's claim that two CSs associate together, but there
is also evidence suggesting, instead, that the second CS links to the UCS.
Group Phase 1 Phase 2
1
loud noise & shock
check for suppression to noise
2
loud noise, light & shock
check for suppression to noise
In Phase 1, we will have exactly the same number of pairings and exactly the same temporal contiguity of the loud noise with the shock. Nevertheless, with rat subjects, we should find considerably smaller conditioned suppression to the noise in Group 2. For this group, there was strong conditioned suppression to the light.
Overshadowing generically refers to a situation in which one element of the compound seems to interfere with the conditioning of another. One way in which to obtain overshadowing is to vary the relative saliences or strengths of the stimuli, as in the experiment above. Alternatively, we can also cause overshadowing by presenting one stimulus more often than another, as in the following design, also based on work by Kamin:
Group Phase 1 Phase 2 Phase 3
1
light, tone & shock (Nothing)
suppression to light?
2
tone & shock
light, tone & shock suppression to light?
In this design, we will find reasonably strong conditioned suppression to the light in Group 1, but not in Group 2. It doesn't necessarily take many conditioning trials to achieve such an effect: In one experiment with rats, Kamin found that 16 pairings of the tone with shock were enough: At that point, additional pairings involving Phase 2 compound conditioning (as in Group 2 above) proved ineffective.
In fact, blocking need not require a procedure in which one CS is presented in an initial phase, and then later compounded with a new CS. Rescorla and Wagner have shown that interspersing compound and simple conditioning trials will do the trick. So, in their experiments, Group 2 above would not have a Phase 1 separate from Phase 2; rather, the tone-&-shock and the tone-light-&-shock trials would be randomly mixed together. What seems to be important here is that one CS is presented more often than another.
Now that you have been introduced to blocking, you may realize that it also can serve to explain a finding we have discussed earlier. That finding is the UCS pre-exposure effect, in which presenting the UCS by itself seems to result in little or no later learning once the UCS is paired with a specific CS. You will recall that one explanation for this effect involved contextual conditioning. Thus, we can now diagram the UCS pre-exposure effect as follows:
Phase 1 Phase 2 Phase 3
Context & UCS Context, CS & UCS CR to CS?
If you compare this to Group 2's procedure above, you will see that, at an abstract level, it is the same.
But what is blocking due to? One possibility is that there is a limit on how much association can form with a single UCS. So, if the association is 'used up' by one CS, then there is nothing left over for another. This account seems unlikely for a number of reasons. One is that the Rescorla and Wagner procedure of interspersing trials would seem to predict that both CSs become conditioned rather than just one, since early in training, the association would not have been all used up. Another is that under certain circumstances, further conditioning is possible, as we will see shortly. Thus, people have looked for other explanations.
Probably the explanation that is most preferred has to do with the notion of signal value (important in the Rescorla and Wagner theory, for example). On that account, to remind you, animals seek to predict future events; they look for good signals that are informative about the UCS. So, a CS that is paired more often with the UCS should have better signal value. To go back to the example of the light, tone, and shock in the Kamin experiment, the tone was present on every trial on which the shock occurred, and was never there when the shock was absent. The tone thus was a perfect predictor of upcoming shock. The light, on the other hand, only predicted a third of the shocks (recall that there were 16 trials with the tone as the only CS, and 8 trials with the tone and light). The tone on this account was clearly a better predictor. If you had your choice regarding what to use as a signal for the presence of crossing cars, and the choice consisted of a red traffic light in front of you that predicted crossing traffic with 95% accuracy (around here, there's often someone who tries to go through a red light!), or someone on the corner with a sign who was only 35% accurate, which signal would be of more value to you?
But the issue here actually goes beyond simple frequencies. In the Kamin and Rescorla-Wagner experiments, there was also redundancy. That is, in these experiments, the light added no new information by itself. Whenever the light was there, so was the tone, making the light perfectly redundant. Accordingly, once the animal has discovered that the tone is the better predictor, and that the tone also predicts in all the situations in which the light occurs, there is no longer any need to pay attention to the light. The animal now has its predictor.
Kamin has made a similar claim; namely, that conditioning will depend on the extent to which the UCS is surprising in a given environment. If it is surprising, then it is unexpected, and the animal will search for a predictor or signal. In this case, finding a good signal can block conditioning with other signals.
Signal value can also account for overshadowing involving differences in salience, of course. More salient CSs are likely to be noticed more, and thus are more likely to be checked out as potential signals or predictors of upcoming biological events.
And finally, signal value can nicely explain a result known as unblocking. If the signal value theory is really correct, then we ought to be able to unblock a stimulus by getting it to predict something new. Or to put this slightly differently, if we make the situation surprising (and not completely predictable) once again, then we ought to jumpstart the search for signals. If our previously blocked CS is now a good signal for the new condition, we ought to find that it has unblocked.
A very nice example of this occurs in an experiment by Dickinson, Hall, and Mackintosh. Here is an overview of part of their experiment:
Group Phase 1 Phase 2
1 light,
2 shocks 4 sec apart light & click, 2 shocks
8 sec apart;
light, 2 shocks 4 sec apart
2
light, 2 shocks 8 sec apart light & click, 2 shocks
8 sec apart;
light, 2 shocks 8 sec apart
In Phase 2, they continued to present the light CS by itself, with the UCS shocks coming in the same pattern they had in Phase 1. But, they also added in compound conditioning. On half of the Phase 2 trials, the light was compounded with a clicking sound.
As you might expect from the discussion so far, the click ought to be blocked in Group 2. It should show no conditioned suppression since it is perfectly redundant with the light, and carries no new information. But the critical comparison involves Group 1. In Phase 2, when the click is presented, it now carries information about how far apart the shocks will be that the light by itself does not. Consistent with this account, the click is unblocked in Group 1, but not Group 2.
Another experiment by Dickinson et al. used the following setup:
Group Phase 1 Phase 2
1
click, 2 shocks 8 sec apart light & click,
2 shocks 8 sec apart
2
click, 2 shocks 8 sec apart light & click,
only 1 shock
From what we know about signal value and redundancy, we should predict the result actually obtained in Group 1: No evidence of conditioning with the light (the light is blocked). However, the light does provide new information in Group 2, since it carries the additional information that there will only be one shock coming up, not two. And in this situation, we find unblocking, as evidenced by aversive conditioning of the light.
A final experiment we can discuss involves returning to an earlier study by Kamin. The setup for this experiment (the last experiment discussed in the overshadowing section above) will now involve one additional group. (Since this group is to be compared with the blocking group, I won't bother re-diagramming the non-blocking Group 1. You can consult the previous section, if you wish, to remind yourself about what that group did.) Thus, for our relevant groups, the methodology involved the following:
Group Phase 1 Phase 2 Phase 3
2
tone & 1 mA shock light,
tone & 1 mA shock suppression
to light?
3
tone & 1 mA shock light,
tone & 4 mA shock suppression
to light?
If you recall our previous discussion of Group 2, you will remember that they block to the light, as it is redundant with the tone. The only additional information I am now giving you about them is how strong a shock (1 mA) they received. But now consider Group 3. Their situation was identical to Group 2's situation, with the exception that the shock was stronger in Phase 2 (it was 4 mA). The change in strength should have been surprising, resulting in unblocking. In fact, the light in this latter condition did unblock, and did act as a conditioned suppressor. So, changing the situation can result in new learning with an otherwise redundant and blocked signal.
Some aspects of blocking can be explained by both Kamin's notion of surprisingness and the Rescorla-Wagner theory. But as you will see in the next chapter, unblocking presents a bit of an embarrassment for the Rescorla-Wagner theory, at least in its mathematical form. Of course, it is also an embarrassment for associational theories that rely strictly on temporal contiguity to explain when conditioning will be successful.
Discrimination Learning Test
CS1 CS2 & UCS
CR to CS1? CR to CS2?
CS2 & no UCS
Normally, in discrimination training, we expect that there should be conditioned inhibition to the CS present in the no-UCS condition (the CS-). That is CS2 . But note that CS2 is also present when the UCS is there. Is it simply ignored, as a strict signal value theory might seem to suggest? (After all, CS1 seems to have better signal value, so far as predicting the UCS is concerned.) Or does its inhibition interfere with the excitation that occurs when the UCS is there? The answer may surprise you.
In the example above, we sometimes find that there is no excitatory CR to CS1 by itself (somewhat contrary to what a strict contingency approach would suggest). Instead, both CSs apparently need to be present for a strong conditioned response. This finding is called occasion setting. One stimulus (CS1 in our example) seems to signal the occasion or circumstances under which another stimulus can be taken as a reliable predictor of the UCS. This result is more often found when the two CSs are separated by an interval of time, but it has been reported in cases involving simultaneous compound conditioning.
Occasion setting provides problems for a number of theories. Most would seem to predict stronger association to CS1 than to CS2 . Indeed, most would seem to predict inhibition to CS2 , so that a sort of summation should occur whereby CS1 and CS2 together result in a smaller CR than when CS1 is presented alone. But that does not happen. Maybe there is a sort of contrast effect influencing what an animal pays attention to here. To remind you, contrasts occur when an animal experiences two levels of a CS. Well, here is a situation where we have something like two levels, in the sense that a single CS is associated with very different outcomes. That should be surprising, in Kamin's sense. And if it is, then perhaps the animal's attention is drawn towards making it less so; that is, towards learning when a CS will be followed by the UCS, and when it won't. In this sense, the occasion setter (CS2 in our example) provides information about CS1, and not the UCS, which may be why the occasion setter by itself does not become excitatory.
As a sample experiment here, consider a study by Balaz, Capra, Hartl, and Miller. Their design involved the following:
Group Phase 1 Phase 2
1
pre-expose unusual context
CS & shock in unusual context
2
(Nothing)
CS & shock in unusual context
You should figure out by now that they are deliberately trying to create latent inhibition for the elements of the unusual context in Group 1. If the context serves as a CS, then pre-exposing it should result in retarded learning for Group 1, compared to Group 2. But, Balaz et al. found that the CS in both groups did indeed become inhibitory. In contrast, contextual cues in Group 1 did not show any evidence of being inhibitory (though from other experiments, they knew that the context cues in Group 2 would be likely to acquire some inhibition once Phase 2 was started). As is typical for occasion setting, the CS in Group 1 had its strongest effect when in the unusual context. Thus, in this series of experiments, we find evidence that context can sometimes serve as a CS (becoming conditioned to the UCS), and at other times may act as an occasion setter.
In our earlier discussion of pre-exposure effects, we found out that contextual conditioning was common; that is, a CS learned in one context or environment did not necessarily trigger a CR in a different context. Within the framework of the current topic, context may be seen to work in part as an occasion setter, so that presence of both the contextual cues and the CS are necessary for the conditioned response.
Another result illustrated in part by this example involves what is called configural conditioning. In configural conditioning, the animal responds to several stimuli in a way that may be completely different from how it responds to each individually. Or to use a phrase made famous by Gestalt psychologists, the whole is greater than the sum of its parts: We cannot account for the reaction to multiple stimuli by adding the reactions to the individual stimuli.
As an illustration, consider the following rather complex discrimination training design:
Discrimination Learning
CS1 & UCS
CS2 & UCS
CS1 CS2 & no UCS
Can we train animals to respond
appropriately? If we train them to give conditioned excitatory responses
to each individual CS, then wouldn't the presence of an excitatory component
in the compound also lead to excitatory responding? In this case,
animals can learn to inhibit the response when both stimuli are
present, and to give it when just one or the other is there. This non-trivial
result is an analogue of the famous XOR problem (for exclusive
or: one or the other, but not both!) in artificial
intelligence (see the chapter on connectionism for more details).
Essentially, Minsky and Pappert used the XOR problem to claim that
a type of artificially intelligent machine called a perceptron would
never be capable of the type of learning humans do. The interesting thing
about a perceptron is that it operates by forming connections or associations
between incoming stimulus components, and outgoing response components.
If this sounds familiar, it is because a perceptron is very much like associationistic
theories making the black box assumption (see Chapter 1). To the
extent that animals can avoid generalization and respond to wholes in completely
different fashion than they respond to the component parts, we seem to
have evidence of an additional layer of representation beyond physical
resemblance, involving complex configural relationships among those elements.
Group CS UCS
1
saccharine water
LiCl injection
2
saccharine water
shock
3
plain water, light, noise
LiCl injection
4
plain water, light, noise
shock
The question, of course, was whether these CSs would become conditioned suppressors. And what they found was this: Groups 1 and 4 showed suppression: Group 1 radically decreased their licking of the saccharine-flavored water, while Group 4 radically decreased their licking of plain water when the light and noise were present. However, Groups 2 and 3 did not show much, if any, conditioned suppression. Such a finding is termed belongingness: It appears to be the case that certain stimuli are more likely to associate with one another than are other stimuli.
Belongingness has been found
in other situations since. But it is a particularly compelling feature
of taste aversions, and has been studied most intensely here. It poses
a major problem for associational theories such as Pavlov's or Watson's
theory, because those theories would seem to predict that any CS and UCS
that are effective should associate with one another (a claim arising from
the general process theory assumption). We know from Group 1 that
the taste of saccharine and the lithium chloride are effective, and we
know from Group 4 that the shock and the light and noise combination are
effective. That would lead us to expect that either UCS ought to result
in conditioning for either CS; but of course, that is not the result Garcia
and Koelling obtained.
Moreover, further striking
a blow against general process theory, a follow-up experiment by Wilcoxin,
Dragoin, and Kral compared belongingness in rats and birds (quail,
specifically). They used an odd-flavored, blue colored water as their CS,
and lithium chloride as their UCS. With rats, the same basic results occurred:
The rats made the connection between taste and getting ill, but they did
not make the connection between visual appearance and getting ill (they
didn't avoid plain-tasting blue water, but they did avoid odd-tasting clear
water). However, the exact opposite pattern occurred for the birds!
Thus, what types of connections are easier to make in part depends on the
species.
 As for why certain species seem to make some connections easier than others,
there are several studies that suggest the explanation may have to do with
what the species is paying attention to when it is eating or drinking.
Smell and taste are important to rats navigating their environments on
foot, whereas visual cues are more likely to be important to birds attempting
to locate prey in mid-flight. Such arguments would suggest that removing
an animal's ability to attend to a certain stimulus dimension might increase
attention to other aspects.
As for why certain species seem to make some connections easier than others,
there are several studies that suggest the explanation may have to do with
what the species is paying attention to when it is eating or drinking.
Smell and taste are important to rats navigating their environments on
foot, whereas visual cues are more likely to be important to birds attempting
to locate prey in mid-flight. Such arguments would suggest that removing
an animal's ability to attend to a certain stimulus dimension might increase
attention to other aspects.
How long a delay can animals tolerate and still show a taste aversion? That was the questioned posed in a study by Kalat and Rozin. They used water flavored with sucrose as their CS, but ran a number of groups of rats that differed in whether the lithium chloride was administered half an hour after the rats tasted the water, or even 6 or 24 hours later! In their test of aversion, they gave their animals two liquids (plain and sweetened water), and tracked how often the animals chose the sweetened water compared to a control group that hadn't gone through associating sweetened water with poison. The results they obtained are presented in Figure 6. Astonishingly, there was still evidence of learning when the CS and the UCS were separated by a full day! Even with that long a delay, the rats with the lithium chloride injection were still not drinking as much of the sweetened water as were the animals who hadn't been put through the taste aversions procedure.
Is there a way to test such an idea? One prediction made by this notion is that long-delay learning ought to be very difficult to obtain for common stimuli, stimuli with which the animal has been familiarized (and thus, in the case of foods, stimuli that can safely be ingested in large quantities or over repeated tastings in a short interval of time). Consistent with this, long-delay learning is more difficult to establish using tastes that presumably have been learned to be safe. Moreover, familiarizing an animal beforehand with an unusual CS (the sucrose or the saccharine -- tastes not found in a rat's natural environment) also reduces the effect. Thus, the results are consistent with specialized learning in the taste system. At the same time, however, it is worth noting that a similar prediction may be generated from the fact that we've pre-exposed the CS.
Finally, consider another interesting experiment on this topic by Kalat and Rozin. They used a setup like the following:
Group Manipulation
1
CS & LiCl 30 minutes later
2
CS & LiCl 240 minutes later
3
CS & LiCl 240 minutes later; CS & 30 LiCl minutes later
A no-brainer prediction here will be that Group 1 will show more aversion than Group 2. But what about Group 3? We might predict from a simple contiguity approach that Group 3 will show the most aversion because they have had two trials of conditioning, and not just one. Moreover, their most recent trial involved the short interval, which we know is generally more effective. In this case, however, Group 3's suppression ratio turned out to be similar to Group 2's ratio. That result is in fact consistent with the learned safety theory: Both Groups 2 and 3 got ill four hours after their first exposure to the poison, so that whatever specialized mechanism is responsible for assessing the safety of a new taste should have reached the same conclusion in both.
A study by Krane and Wagner will give an illustration of this point. Recall that Garcia and Koelling (and many others) found that rats did not associate taste with a shock. Krane and Wagner varied the delay between the CS and the UCS, using values of 5, 30, and 210 sec. They found strong conditioning of a light-noise CS with shock, much as Garcia and Koelling had. But, this association weakened considerably when longer delays were introduced. More interestingly, although they verified the finding that taste and shock did not strongly associate at a 5 sec delay, an association did form with a long delay! Belongingness may thus refer to the optimal time interval required for a given CS and UCS to associate: That time interval may be very different for different stimuli and different species. But that is not quite the same as saying that some associations will not form under any conditions whatsoever.
A similar notion of rehearsal has sometimes been used to account for long-delay learning. Here, an unusual taste is of such potential significance that it is rehearsed or attended for a very long time. In a variant of this theory having to do with distinctiveness, a biologically significant event such as getting ill institutes a special search through memory for any unusual events in the recent past. A novel, distinctive taste will qualify as such an unusual event. When this memory is retrieved, then the memory and the being ill may be rehearsed together, allowing the CS to be connected to the UCS.
The findings briefly reviewed
above are not the only findings found in this interesting paradigm. Another
result (obtained by Bond) that will prove problematic for some associationistic
accounts is that rats who see other rats get sick also start avoiding the
novel taste, even though they were themselves not poisoned. As we will
see, observational learning of this sort places important constraints
on what is necessary for learning to occur in a theory of conditioning.
Is there any compelling evidence in favor of the S-S view? In fact, there is certainly evidence consistent with it. One such piece of evidence involves a phenomenon that we have not yet discussed, sensory preconditioning. Sensory preconditioning explicitly starts off with S-S learning. Probably, the best way to think of sensory preconditioning is that it is the opposite of higher-order conditioning. In higher-order conditioning, we paired two CSs in a second phase; in sensory preconditioning, we reverse the order of the phases. So, the design for sensory preconditioning will be as follows:
Phase 1 Phase 2
CS1 paired with CS2 CS2 paired with UCS
In this paradigm, even though CS1 was never paired with the UCS, and even though there was no discernable CR for either CS at the end of Phase 1, CS1 will, after Phase 2 learning, give the proper conditioned response. One can certainly come up with models here in which the first CS does form a link with the UCS directly, but even these models will have to rely on an association having formed between two supposedly neutral stimuli in Phase 1.
Other evidence that may be interpreted as fitting an S-S view (or at least fitting it better than the S-R view!) involves devaluation studies. In these, following conditioning, there is an attempt to alter the value or meaning of the UCS so as typically to make it less desirable than it was before. In theory, this should not influence the UCR. So, if the CS is directly connected to the UCR, then a devaluation will be irrelevant. But, if the CS is connected to the UCS, then anything affecting that UCS should also affect the ability of the CS to elicit a response. Putting this in terms of correlations, an S-S view would claim that CS and UCS should correlate in the sense that changes to the UCS play out on the CS, since it has to operate through the UCS. But an S-R view claims that the CS and UCS have nothing to do with one another, so that changes to one won't result in corresponding changes with the other.
Let's look at a sample study here. The study should make clear what happens in devaluation. We'll look at Holland and Rescorla. They trained rats using excitatory-appetitive conditioning: a UCS of food coupled with a tone CS. This will normally cause increased activity on the rat's part following the CS. Next in a second phase, they devalued the UCS by essentially briefly spinning rats in a compartment on top of a phonograph (the rats were spun at the rate of 125 rpm). Notice that this happened after ingestion of food (whereas increased activity happens before, and may presumably signal an interest in, the upcoming food). Following the spin treatment, however, the CR of increased activity changed significantly; the animals became a lot less active.
A final example that may be mentioned involves using a response prevention procedure. In this type of experiment, the UCR is prevented from occurring, to see whether learning is still possible. Presumably, if it is, then an association has formed between the CS and the UCS. A sample study here involves Light and Gantt, who temporarily paralyzed a dog's ability to flex its rear leg. They paired a tone with shock to the rear leg. The relevant question was whether, once the dog had recovered, tone would cause the leg flex. It did, supporting an S-S view. (On the other hand, one could argue that the CS center connects to the UCR center; this study did not exclude that possibility, since it involved blocking the motor signals from the brain to the rear paw muscles. That is still a problem for a peripheralist, of course, but not for someone adopting a centralist approach.)
But while there is evidence consistent with the S-S view, there is also evidence that doesn't fit it. Among that evidence is some that we've already mentioned: antagonistic conditioning, for example. Holland adds another piece of evidence to this collection. Holland's design (using rat subjects) was as follows:
Group CS UCS
1
tone Food
2
light Food
As you may remember from
the Holland and Rescorla experiment on devaluation, a tone causes increased
activity in the area where food will be delivered; this is its CR.
A light, on the other hand, causes a very different conditioned response:
significantly decreased movement. On a strict stimulus substitution
approach in which the CS substitutes for the UCS by essentially channeling
excitation or activation to the UCS, both CSs ought to show the same
response. Their failure to do so is incompatible with a simple S-S
model (although it might fit in with more complex models in which the CR
involves aspects of both the reaction to the UCS and the
supposedly neutral reaction to the CS).
In this case, we can trace out the nerve connections. And when we do, we find compelling evidence that the sensory neurons from the gill and tail areas connect (through interneurons) not to one another, but to the motor neuron controlling the gill. That is clearly an S-R connection. Is this how conditioning occurs in other species? Not necessarily. The extraordinary complexity of the nervous system in mammals, for example, makes it difficult to obtain clear evidence for one or the other connection. But as some of the findings above suggest, mammalian conditioning would seem, at least some of the time, to involve S-S rather than just S-R connections.
In the first chapter, I stated that there were likely to be several different learning mechanisms. Here is a case in which we find different but probable mechanisms for classical conditioning. And to complicate the situation even further, note that results with inhibitory techniques, observational learning techniques, and long CS durations (so that the CS turning off is the effective signal) suggest a type of learning in which one of the two events (CS, UCS) is not physically present, a learning that depends on memory representations rather than direct sensory stimulation. The wisest course seems to me to assume that, under the right circumstances, many different types of things can connect, thus accounting for the many different types of effects we obtain.
We have already informally talked about Rescorla's theory: It is a representation-level theory in which learning occurs because the animal acquires new information that increases its ability to predict. If that ability is not modified, then learning will not occur. Thus, the task facing learning theorists is to describe why some arbitrary correlations or co-occurrences (arbitrary in the sense that we set up artificial worlds in a laboratory) appear more effective than others, the problem of determining signal value. But fundamentally, the problem of learning is the problem of information: as information grows, uncertainity decreases. Thus, learning is possible only in situations where there is uncertainity.
With this as background, we may note the extraordinary differences between a definition of classical conditioning like Pavlov's and one like Rescorla's (the learning of relations among events). A Rescorlian animal attempts to predict the future; a Pavlovian animal reacts to the present. A Rescorlian animal is sensitive to cause-and-effect relationships because of the strong correlation between causes and their outcomes. A Pavlovian animal is sensitive to cause-and-effect relationships because causes are normally temporally contiguous to their outcomes. A Rescorlian animal can note absences of stimuli, so that it is capable of tracking negative correlations in which a predictor is informative about absence of some event. It isn't clear how a Pavlovian animal could handle such results outside of extinction or discrimination procedures. And finally, a Rescorlian animal can use its knowledge to prepare for the UCS in an appropriate fashion; it need not be restricted, like the Pavlovian animal, to providing a pale imitation of the UCR. The learning of relations among events suggests the acquisition of a database of information on which to act, a much more active view of behavior and its causes than is to be found in the older associational approaches.
Balaz, M.A., Capra, S., Hartl, P., & Miller, R.R. (1981). Contextual potentiation of acquired behavior after devaluing direct context-US associations. Learning and Motivation, 12, 383-397.
Bolles, R.C. (1970). Species-specific defense reactions and avoidance learning. Psychological Review, 77, 32-48.
Bond, N.W. (1984). The poisoned partner effect in rats: Some parametric considerations. Animal Learning and Behavior, 12, 89-96.
Dickinson, A., Hall, G., & Mackintosh, N.J. (1976). Surprise and the attenuation of blocking. Journal of Experimental Psychology: Animal Behavior Processes, 2, 313-322.
Garcia, J., Ervin, F.R., & Koelling, R.A. (1966). Learning with prolonged delay of reinforcement. Psychonomic Science, 5, 121-122.
Garcia, J., & Koelling, R.A. (1966). Relation of cue to consequence in avoidance learning. Psychonomic Science, 4, 123-124.
Gibbon, J., Baldock, M.D., Locurto, C., Gold, L., & Terrace, H.S. (1977) Trial and intertrial durations in autoshaping. Journal of Experimental Psychology: Animal Behavior Processes, 3, 264-284.
Grice, G.R., & Hunter, (1964).
Harlow, H.F. (1949). The formation of learning sets. Psychological Review, 56, 51-65.
Hebb, D.O. (1949). The organization of behavior: A neuropsychological theory. NY: Wiley.
Hinson, R.E. (1982). Effects of UCS preexposure on excitatory and inhibitory rabbit eyelid conditioning: An associative effect of conditioned contextual stimuli. Journal of Experimental Psychology: Animal Behavior Processes, 8, 49-61.
Hinson, R.E., & Siegel, S. (1980). Trace conditioning as an inhibitory procedure. Animal Learning and Behavior Processes, 8, 49-61.
Holland, P.C., & Rescorla, R.A. (1975). The effect of two ways of devaluing the unconditioned stimulus after first- and second-order appetitive conditioning. Journal of Experimental Psychology: Animal Behavior Processes, 1, 355-363.
Hull, C.L. (1943). Principles of behavior. NY: Appleton-Century-Crofts.
Kalat, J.W. (1977). Status of "learned safety" or "learned noncorrelation" as a mechanism in taste-aversion learning. In L.M. Barker, M.R. Best, & M. Domjan (Eds.), Learning mechanisms in food selection (273-293). TX: Baylor U. Press.
Kalat, J.W., & Rozin, P. (1971). Role of interference in taste-aversion learning. Journal of Comparative and Physiological Psychology, 77, 53-58.
Kalat, J.W., & Rozin, P. (1973). "Learned safety" as a mechanism in long-delay taste-aversion learning in rats. Journal of Comparative and Physiological Psychology, 83, 198-207.
Kamin, L.J. (1965). Temporal and intensity characteristics of the conditioned stimulus. In W.F. Prokasy (Ed.), Classical conditioning: A symposium (118-147). NY: Appleton-Century-Crofts.
Kamin, L.J. (1969). Predictability, surprise, attention, and conditioning. In B.A. Campbell & R.M. Church (Eds.), Punishment and aversive behavior (279-296). NY: Appleton-Century-Crofts.
Krane, R.V., & Wagner, A.R. (1975). Taste aversion learning with a delayed shock US: Implications for the "generality of the laws of learning." Journal of Comparative and Physiological Psychology, 88, 882-889.
Light, J.S., & Gantt, W.H. (1936). Essential part of reflex arc for establishment of conditioned reflex. Formation of conditioned reflex after exclusion of motor peripheral end. Journal of Comparative Psychology, 21, 19-36.
McAllister, W.R. (1953). Eyelid conditioning as a function of the CS-UCS interval. Journal of Experimental Psychology, 45, 417-422.
Minsky, M., & Pappert, S.
Obrist, P.A., Sutterer, J.R., & Howard, J.L. (1972). Preparatory cardiac changes: A psychobiological approach. In A.H. Black & W.F. Prokasy (Eds.), Classical conditioning II: Current theory and research. NY: Appleton-Century-Crofts.
Pavlov, I. (1927). Conditioned reflexes. Oxford: Oxford U. Press.
Rescorla, R.A. (1967). Pavlovian conditioning and its proper control procedures. Psychological Review, 74, 71-80.
Rescorla, R.A., & Wagner, A.R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In A.H. Black & W.F. Prokasy (Eds.), Classical conditioning II: Current theory and research (64-99). NY: Appleton-Century-Crofts.
Rizley, R.C., & Rescorla, R.A.
Schneiderman, N., Fuentes, I., & Gormezano, I. (1962). Acquisition and extinction of the classically conditioned eyelid response in the albino rabbit. Science, 136, 650-652.
Siegel, S. (1972). Conditioning of insulin-induced glycemia. Journal of Comparative and Physiological Psychology, 78, 233-241.
Wagner, A.R. (1981). SOP: A model of automatic memory processing in animal behavior. In N.E. Spear & R.R. Miller (Eds.), Information processing in animals: Memory mechanisms (5-47). NJ: Erlbaum.
Wilcoxin, H.C., Dragoin, W.B., & Kral, P.A. (1971). Illness-induced aversions in rat and quail: Relative salience of visual and gustatory cues. Science, 171, 826-828.
Gone
Fishing: (http://www.mtholyoke.edu/offices/comm/csj/970221/hollis.html)
(a non-technical news release on classical conditioning in fish that you
may enjoy)